



研究内容
生体膜グリセロリン脂質組成は組織や細胞によって異なります。また、グリセロリン脂質は細胞の壁としての役割だけでなく、強力な炎症免疫アレルギーのメディエーターである血小板活性因子(PAF)や呼吸に必須な肺サーファクタント脂質も含みます。極性基、脂肪酸鎖長、脂肪酸の不飽和度を考えると約1000種類の分子種が存在すると考えられています。??この多様性は、リン脂質の代謝回転によって作られており発見者の名からランズ回路(図1)と呼ばれています(Lands, W. E. M. J. Biol. Chem. 1958)。ランズ回路はリン脂質から脂肪酸を遊離させるホスホリパーゼA2 (PLA2)と脂肪酸を再結合させるリゾリン脂質アシル転移酵素によって回転します。これまでPLA2はよく研究されてきましたが、リゾリン脂質アシル転移酵素は膜結合型の酵素であることから、ほとんどが分子同定されず進展が難しい分野でした。
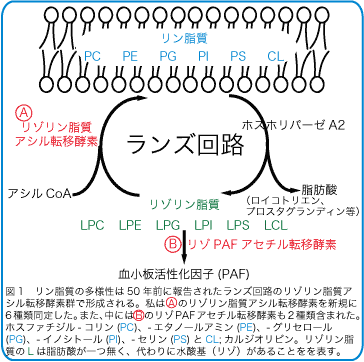
近年、ゲノムデータベースの成熟により、複数のグループから次々とリゾリン脂質アシル転移酵素遺伝子が報告されました。ランズ回路の報告から、実に50年を経ています。2010年5月の時点で11種類のランズ回路のリゾリン脂質アシル転移酵素が報告されており、私達もそのうちの6種類の発見に成功しています(図2赤字)。この分野は今まさにブレイクスルーが起きています。??これらのほとんどは1-acylglycerol-3-phosphate O-acyltransferase (AGPAT)ファミリーかmembrane bound O-acyltransferase (MBOAT)ファミリーに属します。また、これらは複数の活性を持ちます(図2)。
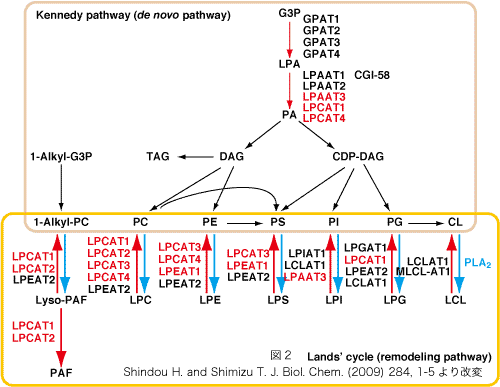
清水研から報告した酵素?LPCAT1(AGPATファミリー)?肺に多く肺サーファクタント脂質産生活性と恒常的なPAF生合成活性を持つ。??LPCAT2(AGPATファミリー)?炎症細胞に発現し、誘導型PAF生合成活性と膜リン脂質(PC)生合成活性を持つ。??LPAAT3(AGPATファミリー)?精巣で週齢依存的に誘導され性成熟に関わると考えられる。PAとPI合成活性??LPCAT3、LPCAT4、LPEAT1(MBOATファミリー)?新規ファミリーを同定した。これらは恒常的に生体膜を合成する酵素だと考えられる。組織分布も基質特性も異なります。ややこしいのですが、LPCAT3はPC、PE、PSをLPCAT4はPC、PEをLPEAT1はPE、PSを生合成します。??他の研究室から報告された酵素を合わせても、まだ未知の酵素は存在すると思います。一つの酵素が複数の基質を認識し、かつ複数の酵素が協調し合い生体膜を作っています(図3)。また、そのバランスが生命維持に重要だと考えています(図4)。
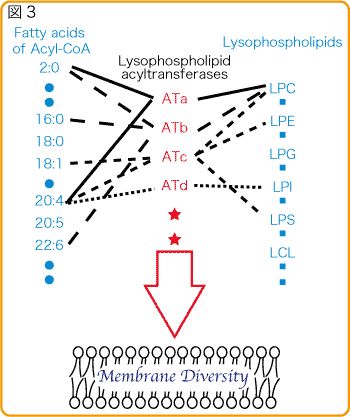
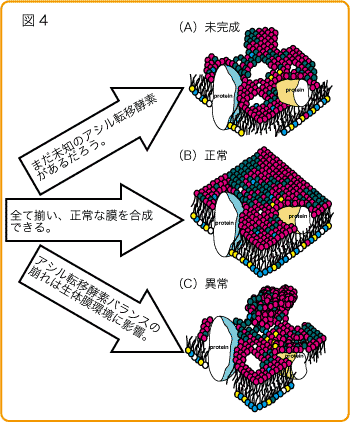
生命が誕生し、進化する上で、この生体膜の役割は大きかったと思われます。様々な細胞機能にも影響を与えているであろう生体膜グリセロリン脂質の多様性形成メカニズムの解明を私達は目指しています。そこから細胞そのものの理解を深め、新たな創薬開発も目指したいと思います。